神経栄養因子
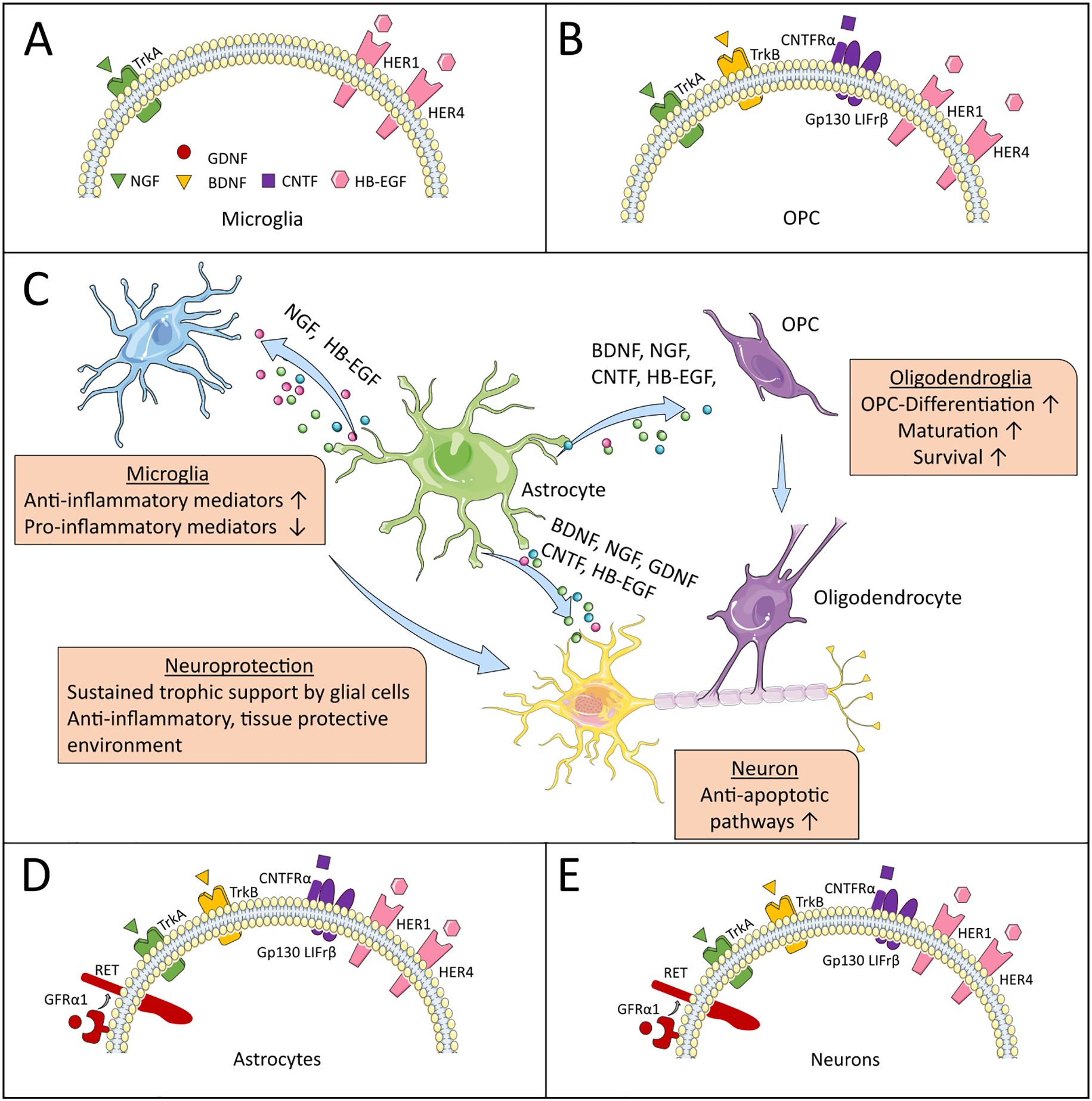
発生期における神経栄養因子の量的制限は、生存するニューロンの数を制御し、ニューロンと適切な密度の標的神経支配の要件との一致を確保します。4つの哺乳類神経栄養因子それぞれの生物学的効果は、トロポミオシン関連キナーゼ(Trk)ファミリーの受容体チロシンキナーゼ(TrkA、TrkB、TrkC)の3つのメンバーの1つまたは複数の活性化を介して媒介されます。さらに、すべての神経栄養因子は、腫瘍壊死因子受容体スーパーファミリーのメンバーであるp75神経栄養因子受容体(p75NTR)を活性化します。Trk受容体への神経栄養因子の結合は、Ras、ホスファチジルイノシトール3-キナーゼ、ホスホリパーゼC-γ1、およびマイトジェン活性化タンパク質キナーゼを含むこれらのタンパク質を介して制御されるシグナル伝達経路の活性化につながります。神経栄養因子の存在は成人期まで必要であり、そこではシナプス機能と可塑性を制御し、ニューロンの生存、形態、分化を維持します。
製品リスト
| ターゲット | カタログ番号 | 製品名 | 反応性 | 予想分子量 |
|---|---|---|---|---|
| Beta-NGF (哺乳類) | PHH1902 | 組換えヒトBeta-NGF(哺乳類) | ヒト | 13.3 KDa |
| NRN1L (C-6His) | PHH1242 | 組換えヒトNRN1L(C-6His) | ヒト | 12.3 KDa |
| CDNF (C-6His) | PHH0389 | 組換えヒトCDNF(C-6His) | ヒト | 19.3 KDa |
| BDNF | PEV0147 | 組換えヒト/マウス/ラットBDNF | ヒト/マウス/ラット | 13 KDa |
| Beta-NGF (110AA) | PEM0151 | 組換えマウスBeta-NGF(110AA) | マウス | 12.4 KDa |
| NT-3 | PEH1860 | 組換えヒトNT-3 | ヒト | 13.6 KDa |
| Beta-NGF | PEH1225 | 組換えヒトBeta-NGF | ヒト | 13.4 KDa |
| pro-Beta NGF | PEH1224 | 組換えヒトpro-Beta NGF | ヒト | 25 KDa |
| Neurturin | PEH1218 | 組換えヒトNeurturin | ヒト | 11.8 KDa |
| Neuritin (N-6His) | PEH1210 | 組換えヒトNeuritin(N-6His) | ヒト | 12.1 KDa |
| GDNF | PEH0723 | 組換えヒトGDNF | ヒト | 15.1 KDa |
| pro-BDNF | PEH0146 | 組換えヒトpro-BDNF | ヒト | 25.6 KDa |
検証データ


関連製品
参考文献
1. 神経栄養因子:概要。Skaper SD。Methods Mol Biol。2018年。[PMID:29222769]
2. 多発性硬化症における神経栄養因子。Güner F、他。Front Immunol。2025年。[PMID:40936917]
