抗体の紹介
抗体の紹介
抗体は、免疫グロブリン(Ig)とも呼ばれ、抗原刺激に応じて免疫系、特にBリンパ球によって産生される高特異性の糖タンパク質のクラスです。適応液性免疫の中核的エフェクター分子であり、病原体感染の防御、異常細胞の除去、体内の恒常性維持において不可欠な役割を果たしています。
I. 基本構造と機能ドメイン
抗体分子の基本構造は、古典的な「Y」字型の4ペプチド鎖対称構造を特徴としています:
-2本の同一な重鎖(Heavy Chain、H鎖):分子量が比較的大きい(約50-75 kDa)。重鎖は抗体のクラスとサブクラスを決定します。
-2本の同一な軽鎖(Light Chain、L鎖):分子量が比較的小さい(約25 kDa)。軽鎖はカッパ(κ)型とラムダ(λ)型に分類されます。
-鎖間ジスルフィド結合:これらの結合は重鎖と軽鎖(H-L)、および2本の重鎖(H-H)を結合し、分子構造の安定性を維持します。
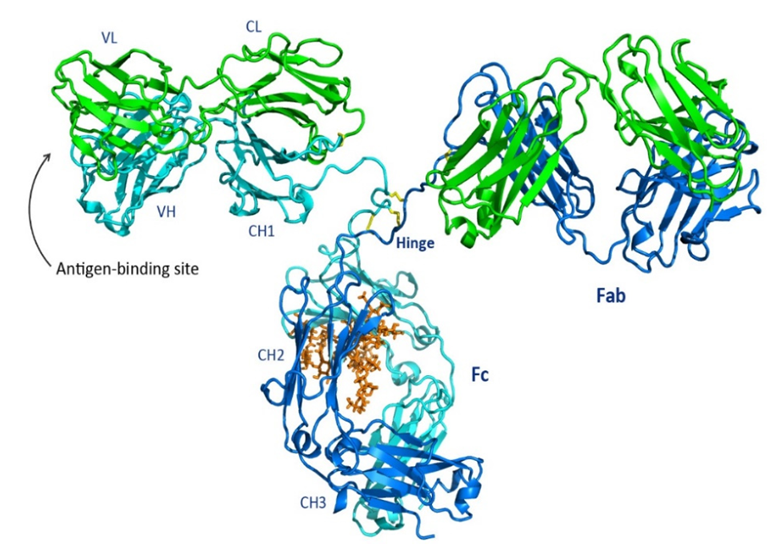
抗体分子はプロテアーゼ(パパインやペプシンなど)によって特徴的なフラグメントに加水分解されます:
-Fabフラグメント(Fragment antigen-binding):完全な軽鎖と重鎖のN末端部分(VHおよびCH1ドメイン)から構成されます。各Fabフラグメントには1つの抗原結合部位があり、抗体の特異性を決定します。この部位は軽鎖と重鎖の可変領域(Variable Region、V領域)によって形成され、高度に可変なアミノ酸配列を持ち、抗原エピトープと直接接触して結合する独特の相補性決定領域(CDR)を形成します。
-Fcフラグメント(Fragment crystallizable):2本の重鎖のC末端部分(IgG、IgA、IgDの場合はCH2およびCH3ドメイン)から構成されます。Fcフラグメントは抗原と結合しませんが、抗体のエフェクター機能を仲介します:
-補体系の結合と活性化(古典経路を介して)。
-Fc受容体を発現する細胞(マクロファージ、NK細胞、好中球、肥満細胞など)と結合し、オプソニン食作用、抗体依存性細胞傷害作用(ADCC)、I型過敏症反応を仲介します。
-胎盤を通過する(IgG)または粘膜表面に輸送される(IgA)。
II. 免疫グロブリンのクラスとサブクラス
重鎖の定常領域(Constant Region、C領域)の抗原性と構造に基づいて、哺乳類の抗体は5つの主要なクラス(アイソタイプ)に分類されます:
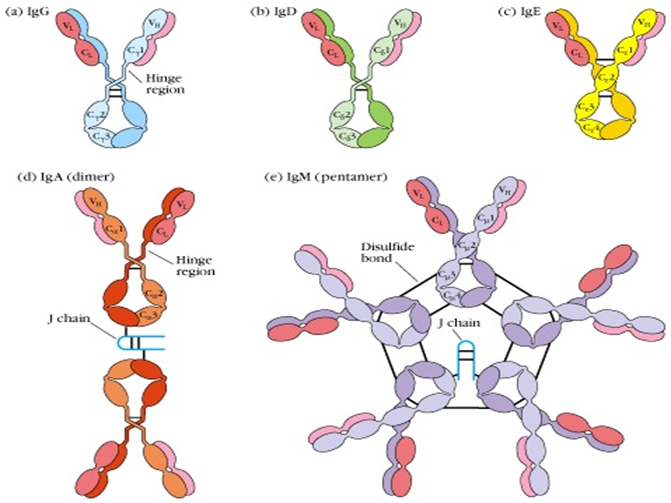
1. IgG:血清中で最も豊富(70-75%)、分子量約150 kDaです。二次免疫応答における主要な抗体であり、強い毒素およびウイルス中和能力を持ち、補体を活性化し、オプソニン作用とADCCを仲介します。胎盤を通過できる唯一の抗体であり、新生児に受動免疫を提供します。ヒトIgGは重鎖の微妙な違いに基づいて4つのサブクラスにさらに分類されます:IgG1、IgG2、IgG3、IgG4。
2. IgM:主に5量体の形で存在(約900 kDa)、一次免疫応答で最初に産生される抗体(「先駆抗体」)です。古典的補体経路を強力に活性化する能力があり(複数のFcセグメントが存在するため)、早期防御、病原体の除去、免疫複合体の処理において重要な役割を果たします。単量体IgMは主にB細胞膜表面に発現し、B細胞受容体(BCR)として機能します。
3. IgA:血清中では主に単量体で存在しますが、粘膜関連リンパ組織(MALT)では多くの場合2量体または3量体を形成し、分泌成分と結合して分泌型IgA(sIgA)を形成します。sIgAは粘膜免疫の主要なバリアであり、唾液、涙、呼吸器および消化管分泌物、初乳中に存在し、病原体の粘膜表面への定着と侵入を防止します。
4. IgD:単量体の形で存在(約180 kDa)、血清中のレベルは極めて低いです。主な機能は成熟B細胞表面の抗原受容体(BCR)の一部として機能し、B細胞の活性化、分化、免疫寛容の維持に関与します。
5. IgE:単量体の形で存在(約190 kDa)、血清中のレベルは最も低いです。そのFcセグメントは肥満細胞や好塩基球表面の高親和性FcεRI受容体に結合します。対応する抗原(アレルゲン)と結合すると、これらの細胞の脱顆粒を引き起こし、ヒスタミンなどの生物活性メディエーターを放出してI型過敏症反応(アレルギー反応)と抗寄生虫免疫を仲介します。
III. 抗体多様性の生成機構
抗体レパートリーはほぼ無限の多様性を持ち、自然界のほぼすべての抗原を認識することができます。この多様性は以下の要因から生じます:
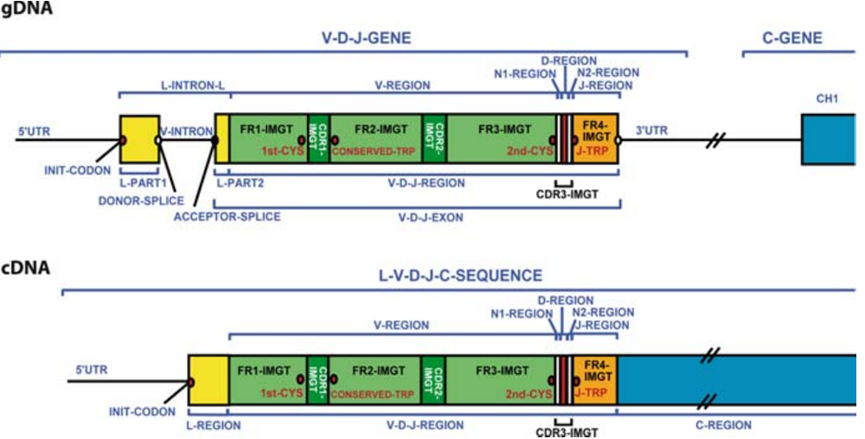
1. 生殖細胞系遺伝子セグメント再配置による組み合わせ多様性:V(D)J遺伝子再配置(重鎖:VH、DH、JHセグメント;軽鎖:VL、JLセグメント)により、多種多様なV領域遺伝子の組み合わせが生成されます。
2. 接合部多様性:V-D、D-J、V-J接合部では、不正確なヌクレオチドの切除と付加(Pヌクレオチド、Nヌクレオチド)により配列変異が生じます。
3. 体細胞超変異:胚中心では、活性化されたB細胞はV領域遺伝子で高頻度の点変異を受け、高親和性抗体の抗原駆動選択を通じて親和性成熟が起こります。
4. 重鎖と軽鎖のランダムな対合:異なる重鎖と軽鎖のランダムな組み合わせにより、さらに多様性が増幅されます。
5. クラススイッチ組換え(CSR):サイトカインやその他のシグナルによって誘導され、B細胞は分泌する抗体のアイソタイプを(IgM/IgDからIgG、IgA、またはIgEに)変更することができます。この際、抗原特異性は変化しません(V領域は不変)、これにより多様な免疫ニーズを満たすために異なるエフェクター機能を獲得します。
IV. 抗体の中核機能
1. 抗原の特異的認識と結合:これは抗体の最も基本的な機能です。V領域のCDRを通じて、抗体は抗原上の特定のエピトープ(病原体表面タンパク質、毒素、外因性高分子など)を精密に認識し結合します。
2. 中和:抗体は病原体(ウイルス、細菌毒素など)のキー部位に結合し、宿主細胞受容体との相互作用や細胞への侵入を阻止することで、それらの感染性や毒性を中和します。
3. オプソニン作用:抗体(主にIgG)のFabセグメントが粒子抗原(病原体など)に結合し、Fcセグメントが食細胞(マクロファージ、好中球)上のFcγRに結合することで、これらの細胞の食作用効率が大幅に向上します。
4. 補体活性化:抗原に結合した抗体(主にIgMとIgG1/IgG3)は免疫複合体を形成し、補体系の古典経路を効果的に活性化し、様々な効果をもたらします:膜侵襲複合体(MAC)の形成による病原体や細胞の溶解(溶解作用)、オプソニン(C3b)の生成による食作用の増強、走化性因子(C3a、C5a)の生成による炎症細胞の遊走。
5. 抗体依存性細胞傷害作用(ADCC):抗体(主にIgG)のFabセグメントが標的細胞(ウイルス感染細胞や腫瘍細胞など)上の抗原に結合し、Fcセグメントがエフェクター細胞(NK細胞など)上のFcγRIII(CD16)に結合することで、これらのエフェクター細胞を活性化し、パーフォリンやグランザイムなどの細胞傷害性分子を放出して標的細胞を殺傷します。
6. 過敏症反応の仲介:IgEは肥満細胞/好塩基球上のFcεRIに結合し、アレルゲンによる架橋結合が起こると、脱顆粒とI型過敏症反応が引き起こされます。特定のIgG/IgMもII型(細胞毒性)およびIII型(免疫複合体媒介)の過敏症反応に関与することがあります。
7. 免疫調節:抗体はイディオタイプネットワークを介して免疫応答を調節することができます。B細胞上のFcγRIIBにFcセグメントが結合すると抑制シグナルが伝達され、B細胞の活性が低下します。抗体フィードバック阻害も液性免疫を調節する重要なメカニズムです。
抗体は免疫系の精緻で強力な兵器庫です。その巧妙なY字型構造、特に高度に可変な抗原結合フラグメント(Fab)と定常エフェクター領域(Fc)は、ほぼ無限の多様性の抗原を認識し、多様な免疫機能を発揮する能力を抗体に与えています。研究ツールとして、タンパク質精製、細胞または組織の局在化研究(WB、IHC、IFなど)、機能研究において重要な役割を果たしています。
EnkiLifeでは、幅広い研究ニーズに応えるために、高品質な抗体を多数取り揃えています。抗体カタログを検索してください。
モノクローナル抗体技術の成熟と抗体工学の急速な発展に伴い、抗体は基礎免疫学研究の対象から現代医療診断、予防、治療の礎として進化しました。がんや自己免疫疾患などの主要疾患領域で革命的な治療効果を示し、生物医学イノベーションの最前線をリードし続けています。
 | Dylan Z Dylan ZはEnkiLifeのタンパク質・抗体専門家で、タンパク質発現システムと抗体調製技術に精通しています。技術において卓越性を追求し、安定で使いやすい製品をユーザーに提供することに尽力しています。 |