cAMPタンパク質キナーゼ触媒サブユニットウサギモノクローナル抗体
コンジュゲーション: 非共役
組換えウサギモノクローナル抗体
アプリケーション
反応性
ヒト、マウス、ラット
遺伝子名
cAMP Protein Kinase Catalytic Subunit
保存
小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。
要約
| 製品名 | cAMPタンパク質キナーゼ触媒サブユニットウサギモノクローナル抗体 |
| 説明 | 組換えウサギモノクローナル抗体 |
| 宿主 | うさぎ |
| 反応性 | ヒト、マウス、ラット |
| コンジュゲーション | 非共役 |
| 修飾 | 未修正 |
| アイソタイプ | IgG |
| クローン性 | モノクローナル |
| 形態 | 液体 |
| 濃度 | 非共役 |
| 保存 | 小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。 |
| 配送 | 氷嚢。 |
| バッファー | 0.05% アジ化ナトリウム、0.05% 保護タンパク質、50% グリセロールを含む TBS で精製された抗体。 |
| 精製 | アフィニティー精製 |
抗原情報
| 遺伝子名 | cAMP Protein Kinase Catalytic Subunit |
| 別名 | PKACA; PPNAD4 |
| 遺伝子ID | 5566 |
| SwissProt ID | P17612 |
| 免疫原 | ヒトcAMPタンパク質キナーゼ触媒サブユニットの合成ペプチド |
アプリケーション
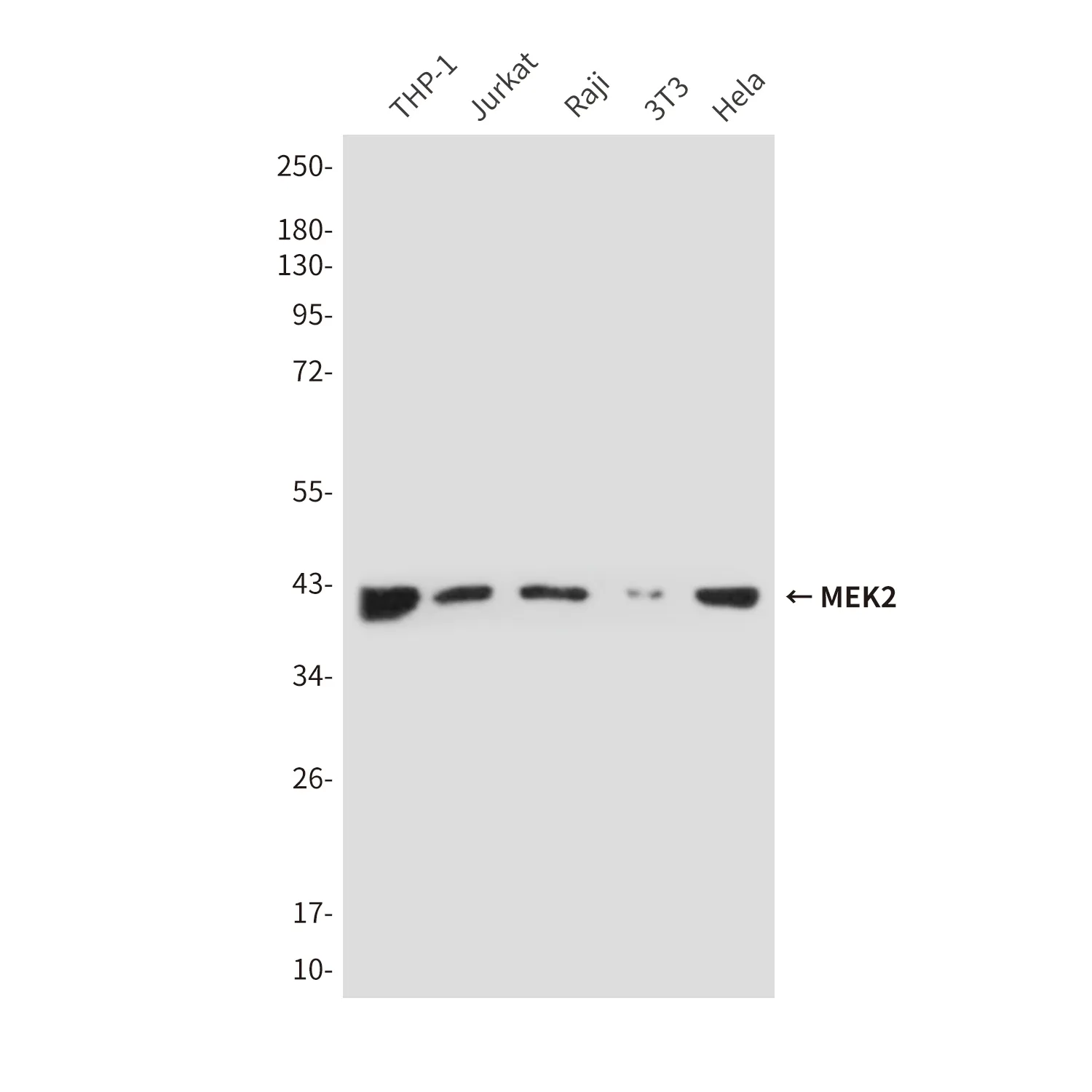
| アプリケーション | WB,IHC,ICC,IP |
| 希釈倍率 | WB 1:500-1:1000,IHC 1:50-1:100,ICC 1:50-1:200,IP 1:10-1:20 |
| 分子量 | Calculated MW: 41 kDa; Observed MW: 41 kDa |
研究分野
| MAPK signaling pathway |
背景
| 細胞質および核内の多数の基質をリン酸化します。PJA2のリン酸化を介して、制御サブユニットの区画化されたプールの存在量を調節します。PJA2はこれらのサブユニットに結合し、ユビキチン化してタンパク質分解を引き起こします。CDC25B、ABL1、NFKB1、CLDN3、PSMC5/RPT6、PJA2、RYR2、RORA、VASPをリン酸化します。RORAはリン酸化によって活性化されます。骨芽細胞からのグルコース媒介性脂肪分化の促進と骨分化の阻害に必要です。トロンビンおよびコラーゲンに対する血小板の調節に関与します。 NF-κB (NFKB1 および NFKB2) および I-κB-α (NFKBIA) と複合した際に、多数の血小板阻害経路のタンパク質をリン酸化することにより、循環血小板を休止状態に維持しますが、トロンビンとコラーゲンがこれらの複合体を破壊し、遊離活性 PRKACA が血小板を刺激し、VASP をリン酸化して血小板凝集を引き起こします。活性化されると、乳がん細胞におけるα-ジフルオロメチルオルニチンの抗増殖効果と抗浸潤効果を阻害します。RYR2 チャネルの活性は、管腔内 Ca2+ の存在下でのリン酸化によって増強され、ストア過負荷誘発性 Ca2+ 放出 (SOICR) の振幅の減少と頻度の増加をもたらします。SOICR は、波の振幅の減少と細胞質 Ca2+ の休止にもかかわらず、Ca2+ 放出速度と自発的な Ca2+ 波の伝播速度の増加を特徴とします。リン酸化による PSMC5/RPT6 の活性化は、プロテアソームを刺激します。 CLDN3 のリン酸化を介して卵巣がん細胞のタイトジャンクション (TJ) を負に制御します。NFKB1 のリン酸化は NF-κB p50-p50 DNA 結合を促進します。胚のパターン形成と形態形成を決定するヘッジホッグ (Hh) シグナル伝達経路を下方制御することで胚発生に関与します。リン酸化による CDC25B の不活性化を介して、前期停止卵母細胞での減数分裂再開を阻止します。また、脚橋被蓋核 (PPT) での急速眼球運動 (REM) 睡眠も制御する可能性があります。APOBEC3G と AICDA をリン酸化します。アイソフォーム 2 は精子鞭毛の ABL1 をリン酸化して活性化し、精子の受精能獲得を促進します。HSF1 をリン酸化します。このリン酸化は熱ショック時のHSF1の核局在と転写活性を促進します(PubMed:21085490)。 |