TMS1 (2N17) ウサギモノクローナル抗体
コンジュゲーション: 非共役
組換えウサギモノクローナル抗体
アプリケーション
反応性
人間
遺伝子名
PYCARD
保存
小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。
要約
| 製品名 | TMS1 (2N17) ウサギモノクローナル抗体 |
| 説明 | 組換えウサギモノクローナル抗体 |
| 宿主 | うさぎ |
| 反応性 | 人間 |
| コンジュゲーション | 非共役 |
| 修飾 | 未修正 |
| アイソタイプ | IgG |
| クローン性 | モノクローナル |
| 形態 | 液体 |
| 濃度 | 非共役 |
| 保存 | 小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。 |
| 配送 | 氷嚢。 |
| バッファー | ウサギIgG(リン酸緩衝生理食塩水、pH 7.4、150mM NaCl、0.02%新型保存料N、50%グリセロール含有)。短期保存は+4℃、長期保存は-20℃で保存してください。凍結融解サイクルは避けてください。 |
| 精製 | アフィニティー精製 |
抗原情報
| 遺伝子名 | PYCARD |
| 別名 | PYCARD; ASC; CARD5; TMS; TMS-1; TMS1; |
| 遺伝子ID | 29108 |
| SwissProt ID | Q9ULZ3 |
| 免疫原 | ヒトTMS1の合成ペプチド |
アプリケーション
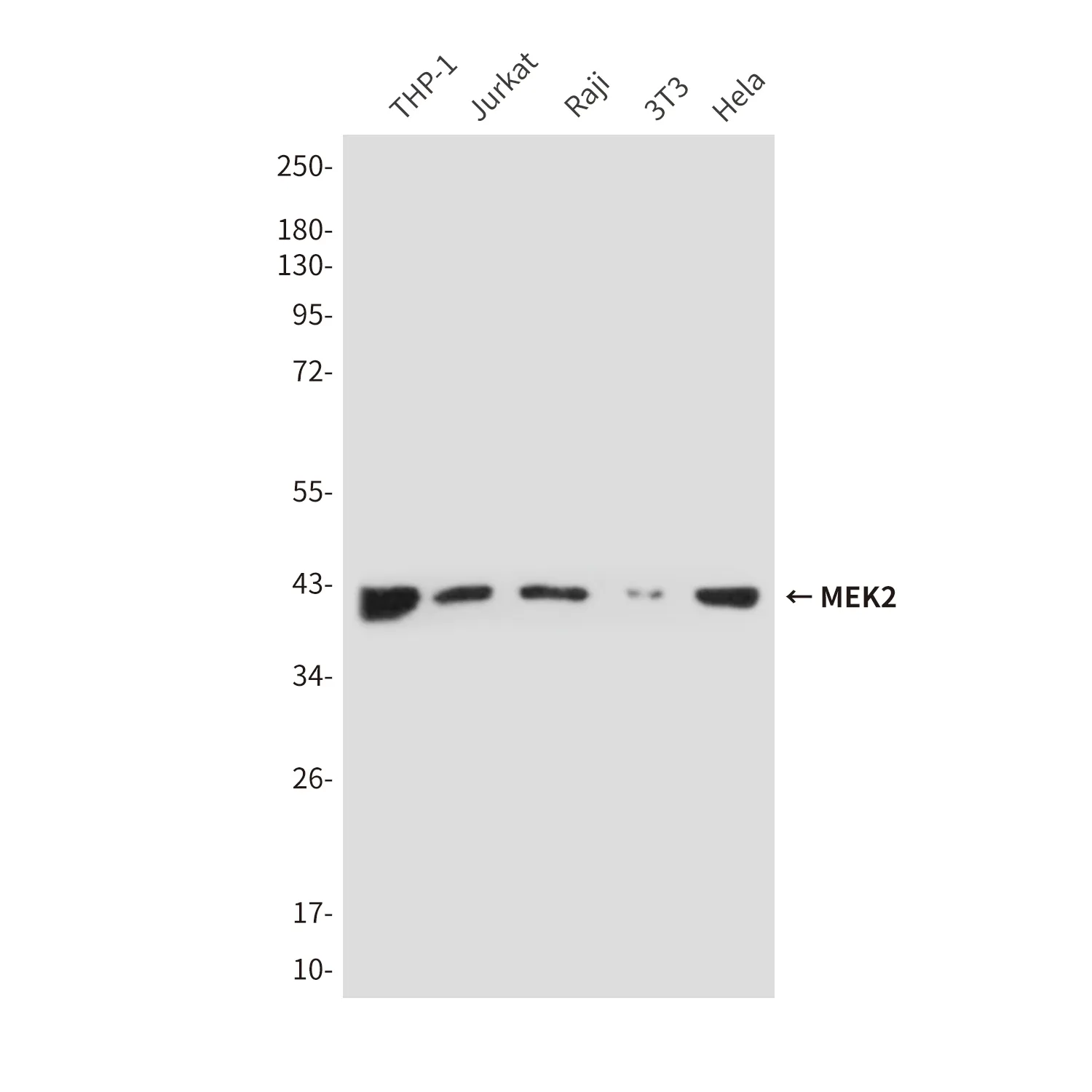
| アプリケーション | WB,ICC/IF,FC |
| 希釈倍率 | WB 1:1000-1:5000,ICC/IF 1:100-1:200,FC 1:10-1:100 |
| 分子量 | 22kDa |
研究分野
| Cell Biology |
背景
| TMS1(メチル化誘導サイレンシングの標的)/ASC(CARDを含むアポトーシス関連スペックルタンパク質)は、PYCARDおよびCARD5とも呼ばれ、N末端ピリンドメイン(PYD)およびC末端カスパーゼリクルートメントドメイン(CARD)を含む22kDaのプロアポトーシスタンパク質です。アポトーシスおよび炎症における重要なメディエーターとして機能します。主にカスパーゼ8、またおそらくはカスパーゼ9が関与するカスパーゼ介在アポトーシスを、おそらく細胞タイプ特異的な方法で促進します。ミトコンドリアアポトーシス経路の活性化に関与し、特定の細胞タイプにおいてFADDとは独立してカスパーゼ8依存性BIDのタンパク質分解成熟を促進し、さらにBAXのミトコンドリア移行を媒介し、カスパーゼ9、2、および3の活性化と結合したBAX依存性アポトーシスを活性化します。マクロファージのピロプトーシス(カスパーゼ1依存性の炎症性細胞死)に関与し、カリウム枯渇時に形成され、カスパーゼ1を急速にリクルートして活性化するASCピロプトソームの主成分である。自然免疫応答では、インフラマソームの組み立てにおいて不可欠なアダプターとして作用し、カスパーゼ1を活性化して炎症性サイトカインの処理と分泌につながると考えられている。さまざまな種類のインフラマソームにおける活性化アダプターとしての機能は、ピリンおよびCARDドメインとそれらのホモタイプ相互作用によって媒介される。NLRP2、NLRP3、AIM2、おそらくIFI16などの特定のパターン認識受容体を含むインフラマソームへのカスパーゼ1のリクルートに必要。NLRP1およびNLRC4インフラマソームでは、必須ではないようだが、プロカスパーゼ1の処理を促進する。 NOD2 と連携して、細菌のムラミルジペプチドによって活性化されたインフラマソームに関与し、カスパーゼ 1 の活性化につながる。DDX58 が引き起こす炎症誘発反応およびインフラマソームの活性化に関与している可能性がある。アイソフォーム 2 は、インフラマソーム アダプターとしての機能に制御効果を持つ可能性がある。アイソフォーム 3 は、インフラマソームを介したインターロイキン-1 ベータの成熟を阻害すると思われる。細胞質二本鎖 DNA を検出する AIM2 と連携して、カスパーゼ 8 が関与するカスパーゼ 1 非依存性細胞死にも関与している可能性がある。適応免疫では、樹状細胞の成熟に関与して T 細胞免疫を刺激し、走化性および抗原取り込みと結びついた細胞骨格の再編成では、グアニンヌクレオチド交換因子 DOCK2 の転写後制御に関与している可能性がある。後者の機能は核型に関与することが提案されている。インフラマソームとは独立してサイトカインおよびケモカインの転写活性化にも関与し、この機能にはAP-1、NF-κB、MAPK、およびカスパーゼ8シグナル伝達経路が関与している可能性がある。NF-κB活性化および阻害機能の調節については、報告されている。CHUKおよびIKBKのキナーゼ活性を阻害することにより、IKK複合体レベルでNF-κB誘導を調節する。CASP1との会合においてRIPK2と競合し、CASP1を介したRIPK2依存性NF-κB活性化をダウンレギュレーションし、インターロイキン-1βプロセシングを活性化することが示唆されている。細胞質二本鎖DNAの存在下でCGASの切断を誘導し不活性化することにより、DNAウイルス感染に対する宿主の抵抗性を調節すると考えられる(PubMed:28314590)。 |