SGK1 (12S19) ウサギモノクローナル抗体
コンジュゲーション: 非共役
組換えウサギモノクローナル抗体
アプリケーション
反応性
ヒト、マウス、ラット
遺伝子名
SGK1
保存
小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。
要約
| 製品名 | SGK1 (12S19) ウサギモノクローナル抗体 |
| 説明 | 組換えウサギモノクローナル抗体 |
| 宿主 | うさぎ |
| 反応性 | ヒト、マウス、ラット |
| コンジュゲーション | 非共役 |
| 修飾 | 未修正 |
| アイソタイプ | IgG |
| クローン性 | モノクローナル |
| 形態 | 液体 |
| 濃度 | 非共役 |
| 保存 | 小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。 |
| 配送 | 氷嚢。 |
| バッファー | ウサギIgG(リン酸緩衝生理食塩水、pH 7.4、150mM NaCl、0.02%新型保存料N、50%グリセロール含有)。短期保存は+4℃、長期保存は-20℃で保存してください。凍結融解サイクルは避けてください。 |
| 精製 | アフィニティー精製 |
抗原情報
| 遺伝子名 | SGK1 |
| 別名 | EC 2.7.11.1; SGK; SGK1; Serum/glucocorticoid-regulated kinase; Sgk; Sgk1; kinase SGK; |
| 遺伝子ID | 6446 |
| SwissProt ID | O00141 |
| 免疫原 | ヒトSGK1の合成ペプチド |
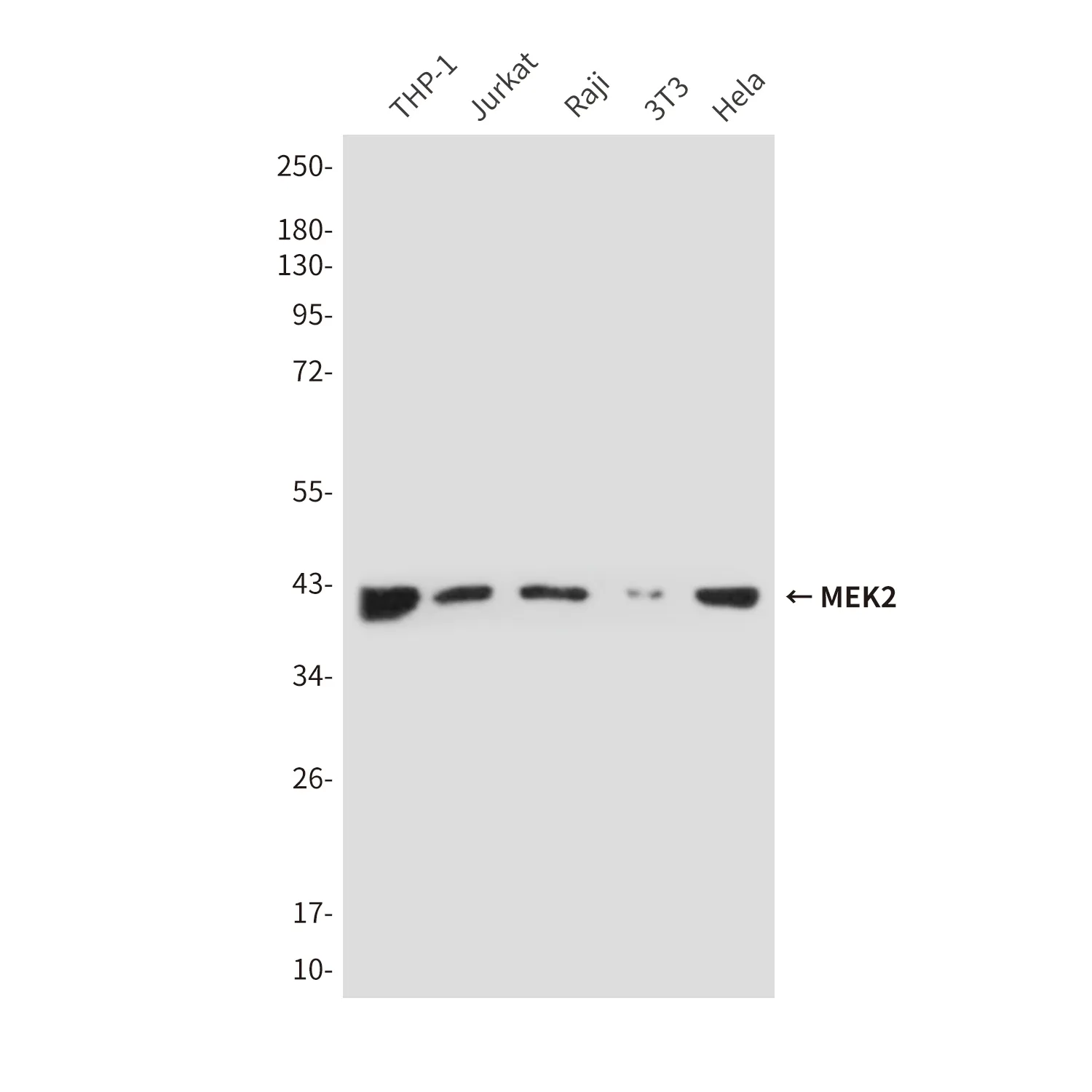
アプリケーション
| アプリケーション | WB,IHC,FC,IP,IF-P |
| 希釈倍率 | WB 1:500-1:2000,IHC 1:50-1:200,FC 1:20-1:50,IP 1:20-1:50,IF-P 1:50-1:200 |
| 分子量 | 49kDa |
研究分野
| Signal Transduction |
背景
| 血清およびグルココルチコイド誘導性キナーゼ(SGK)は、Aktと密接な関連のあるセリン/スレオニンキナーゼです。SGKは、血清、グルココルチコイド、卵胞刺激ホルモン、浸透圧ショック、ミネラルコルチコイドなど、さまざまな刺激に反応して急速に誘導されます。SGKの活性化は、HGF PI3K依存性経路、およびインテグリンを介したPI3K非依存性経路によって達成されます。SGKの誘導と活性化は、抗アポトーシスおよび細胞周期制御の調整を活性化することに関係していると考えられています。セリン/スレオニンタンパク質キナーゼは、さまざまなイオンチャネル、膜輸送体、細胞酵素、転写因子、神経興奮性、細胞成長、増殖、生存、遊走、アポトーシスの制御に関与しています。細胞ストレス応答において重要な役割を果たします。腎臓のNa(+)保持、腎臓のK(+)排泄、塩分欲求、胃酸分泌、腸管のNa(+)/H(+)交換および栄養素輸送、血圧のインスリン依存性塩感受性、末梢グルコース取り込みの塩感受性、心臓の再分極および記憶の強化の調節に寄与します。 Na(+) チャネルをアップレギュレート: SCNN1A/ENAC、SCN5A、ASIC1/ACCN2、K(+) チャネル: KCNJ1/ROMK1、KCNA1-5、KCNQ1-5、KCNE1、上皮 Ca(2+) チャネル: TRPV5 および TRPV6、塩素チャネル: BSND、CLCN2、CFTR、グルタミン酸トランスポーター: SLC1A3/EAAT1、SLC1A2 /EAAT2、SLC1A1/EAAT3、SLC1A6/EAAT4、SLC1A7/EAAT5、アミノ酸トランスポーター: SLC1A5/ASCT2、SLC38A1/SN1、SLC6A19、クレアチントランスポーター: SLC6A8、Na(+)/ジカルボキシレート共輸送体: SLC13A2/NADC1、Na(+) 依存性リン酸共輸送体をアップレギュレート: SLC34A2/NAPI-2B、グルタミン酸受容体:GRIK2/GLUR6。キャリア:SLC9A3/NHE3、SLC12A1/NKCC2、SLC12A3/NCC、SLC5A3/SMIT、SLC2A1/GLUT1、SLC5A1/SGLT1、SLC15A2/PEPT2をアップレギュレーションする。酵素:GSK3A/B、PMM2、Na(+)/K(+) ATPase、転写因子:CTNNB1、核因子NF-κBを調節する。SCNN1A/ENACの安定性と発現を高めることで、上皮細胞へのナトリウム輸送を促進する。これは、NEDD4LユビキチンE3リガーゼをリン酸化することで達成され、14-3-3タンパク質との相互作用を促進することで、SCNN1A/ENACへの結合を阻害し、分解の標的とします。 ORAI1およびSTIM1を刺激してストア作動性Ca(+2)流入(SOCE)を制御する。KCNJ1/ROMK1をリン酸化によって直接制御するか、SLC9A3R2/NHERF2との相互作用の増加によって間接的に制御する。MDM2をリン酸化して、MDM2依存性のp53/TP53ユビキチン化を活性化する。MAPT/TAUをリン酸化して、海馬ニューロンにおける微小管脱重合および神経突起形成を媒介する。SLC2A4/GLUT4をリン酸化して、その活性をアップレギュレーションする。APBB1/FE65をリン酸化して、その核局在を促進する。MAPK1/ERK2をリン酸化して、MAP2K1/MEK1およびMAP2K2/MEK2との相互作用を強化することで活性化する。FBXW7をリン酸化して、NOTCH1シグナル伝達において阻害的な役割を果たす。 FOXO1をリン酸化することで、核から細胞質への再局在化を誘導する。FOXO3をリン酸化することで、核からの脱却を促進し、FOXO3依存性転写を阻害する。BRAFおよびMAP3K3/MEKK3をリン酸化して、それらの活性を阻害する。デキサメタゾンに反応してSLC9A3/NHE3をリン酸化することで、その活性化と細胞膜への局在化を促進する。CREB1をリン酸化。血管新生における血管リモデリングに必要。高レベルおよび高活性が持続すると、高血圧や糖尿病性腎症などの病態に寄与する可能性がある。アイソフォーム2は、アイソフォーム1よりもSCNN1A/ENACの細胞膜発現およびNa(+)輸送に大きな影響を示した。 |