IGF2BP1ウサギモノクローナル抗体
コンジュゲーション: 非共役
組換えウサギモノクローナル抗体
アプリケーション
反応性
ヒト、マウス
遺伝子名
IGF2BP1
保存
小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。
要約
| 製品名 | IGF2BP1ウサギモノクローナル抗体 |
| 説明 | 組換えウサギモノクローナル抗体 |
| 宿主 | うさぎ |
| 反応性 | ヒト、マウス |
| コンジュゲーション | 非共役 |
| 修飾 | 未修正 |
| アイソタイプ | IgG |
| クローン性 | モノクローナル |
| 形態 | 液体 |
| 濃度 | 非共役 |
| 保存 | 小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。 |
| 配送 | 氷嚢。 |
| バッファー | 50mMトリスグリシン(pH 7.4)、0.15M NaCl、40%グリセロール、0.01%アジ化ナトリウム、0.05%保護タンパク質 |
| 精製 | アフィニティー精製 |
抗原情報
| 遺伝子名 | IGF2BP1 |
| 別名 | insulin-like growth factor 2 mRNA binding protein 1; IMP1; ZBP1; CRDBP; IMP-1; CRD-BP; VICKZ1 |
| 遺伝子ID | 10642 |
| SwissProt ID | Q9NZI8 |
| 免疫原 | ヒトIGF2BP1の組換えタンパク質 |
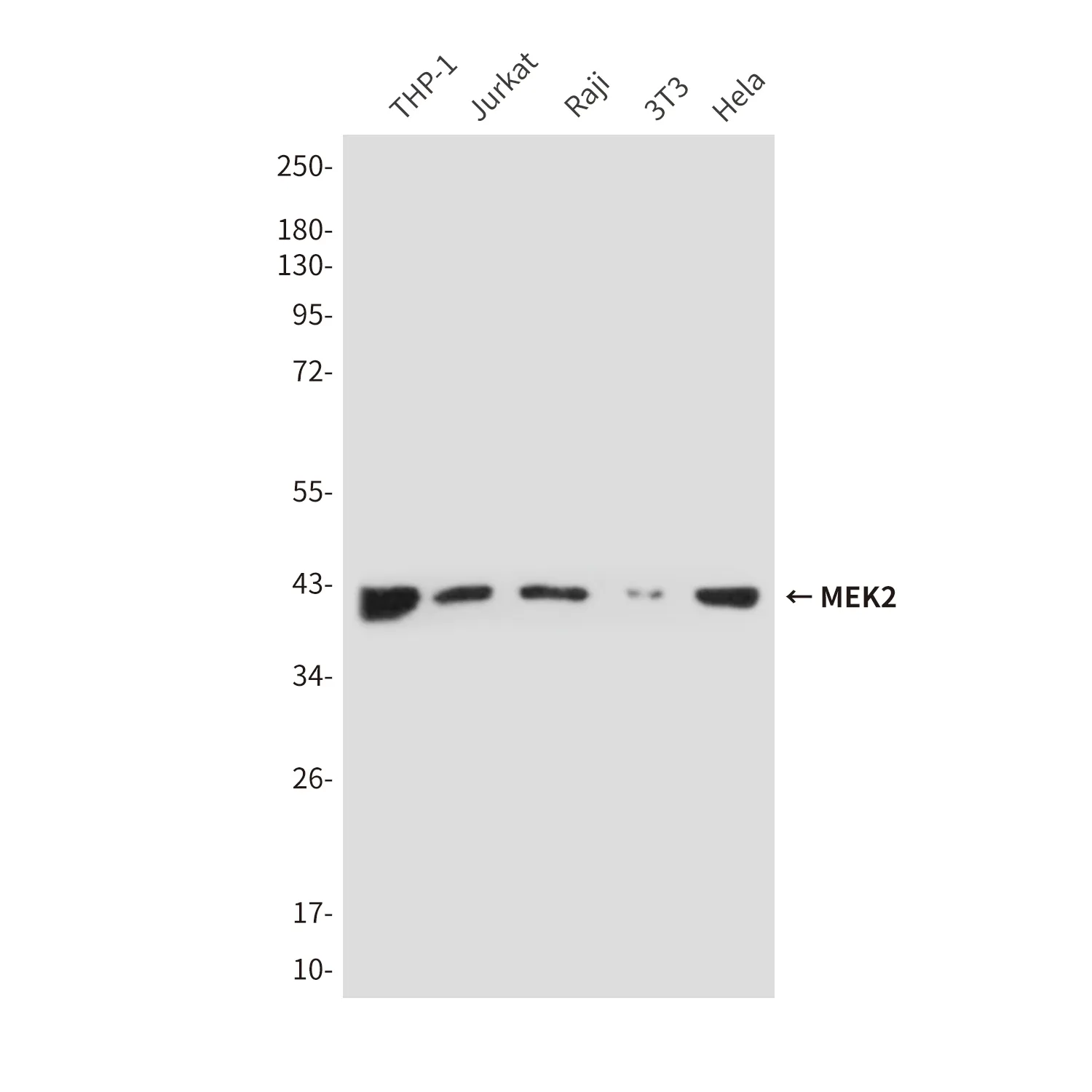
アプリケーション
| アプリケーション | WB,IHC,IP |
| 希釈倍率 | WB 1:500-1:1000,IHC 1:50-1:100,IP 1:20-1:50 |
| 分子量 | Calculated MW: 63 kDa; Observed MW: 63 kDa |
研究分野
| Epigenetics and Nuclear Signaling |
背景
| 標的転写産物を細胞質タンパク質-RNA複合体(mRNP)にリクルートするRNA結合因子。この転写産物のmRNPへの「ケージング」により、mRNAの輸送と一時的な保管が可能になる。また、標的転写産物が翻訳装置に遭遇する速度と位置を調節し、エンドヌクレアーゼ攻撃やマイクロRNAを介した分解から保護する。成体感覚ニューロンにおける軸索再生に必要な転写産物の輸送と翻訳に直接関与する。細胞極性、細胞移動、神経突起伸展に重要なプロセスである、局所的なβアクチン/ACTB mRNAの翻訳を制御する。核内でACTB mRNAと共転写的に会合する。この結合には、ACTB mRNA 3'-UTR内の「ジップコード」として知られる54ヌクレオチドの保存された要素が関与する。こうして形成されたRNPは細胞質へ輸送され、モータータンパク質に結合し、細胞骨格に沿って細胞周縁部へ輸送される。輸送中、ACTB mRNAがタンパク質に翻訳されるのを防ぐ。RNP複合体が細胞膜付近の目的地に到達すると、IGF2BP1がリン酸化される。これによりmRNAが放出され、リボソーム40Sおよび60Sサブユニットが組み立てられ、ACTBタンパク質合成が開始される。次に単量体ACTBは皮質下アクチン細胞骨格に組み立てられる。ニューロンの発達中、神経突起伸展、成長円錐ガイダンス、およびニューロン細胞の移動の主要な制御因子であり、おそらくACTBなどのタンパク質合成の時空間的微調整を介している。活性化シナプスへのmRNA輸送を制御している可能性がある。ABCB1/MDR-1 mRNAに結合して安定化する。間質創傷修復中、PTGS2転写産物と相互作用して安定化する。 PTGS2 mRNAの安定化は、結腸粘膜創傷治癒に非常に重要であると考えられる。協調的かつ逐次的な二量体形成のメカニズムによってIGF2 mRNAの3'-UTRに結合し、IGF2 mRNAの細胞内局在および翻訳を制御する。オープンリーディングフレーム(ORF)のコード領域不安定性決定因子(CRD)内のMYC mRNAに結合し、エンドヌクレアーゼによるMYC切断、およびおそらくMYC-CRDを標的とするマイクロRNAを防ぐ。CD44 mRNAの3'-UTRに結合して安定化し、癌細胞における細胞接着および浸潤突起形成を促進する。癌胎児性H19転写産物およびニューロン特異的TAU mRNAに結合し、それらの局在を制御する。BTRC/FBW1A mRNAに結合して安定化する。 PABPC1 mRNAに存在するアデニンリッチ自己調節配列(ARS)に結合し、その翻訳を抑制します。PABPC1 mRNAへの結合はPABPC1タンパク質によって刺激されます。AGO2とのマイクロRNA依存性相互作用を阻害することにより、BTRC/FBW1A mRNAの分解を阻害します。細胞内シグナル伝達ネットワークを微調整することにより、腫瘍由来細胞の方向性のある移動を促進します。MAPK4 3'-UTRに結合し、その翻訳を阻害します。PTEN転写産物のオープンリーディングフレーム(ORF)と相互作用し、mRNAの分解を阻害します。MAPK4(ダウンレギュレーション)とPTEN(アップレギュレーション)に対するこの複合作用は、HSPB1のリン酸化を拮抗させ、結果としてリン酸化HSPB1によるGアクチンの隔離を阻害し、Fアクチンの重合を可能にします。したがって、細胞移動速度が向上し、PTEN調節による分極によって方向性のある細胞移動が刺激されます。 C型肝炎ウイルス(HCV)の5'-UTRおよび3'-UTRと相互作用し、HCV IRESにおける翻訳を特異的に促進します。ただし、5'-キャップ依存性翻訳は促進しません。これはおそらくeIF3をリクルートすることによって促進されます。HIV-1 GAGタンパク質と相互作用し、感染性HIV-1粒子の形成を阻害します。ウイルスRNAのパッケージング、および細胞膜上でのGAGタンパク質の組み立てと処理を阻害することで、HIV-1の組み立てを抑制します。酸化ストレスや熱ショックなどの細胞ストレス時には、ストレス顆粒にリクルートされる標的mRNA(CD44、IGF2、MAPK4、MYC、PTEN、RAPGEF2、RPS6KA5転写産物など)を安定化します。 |