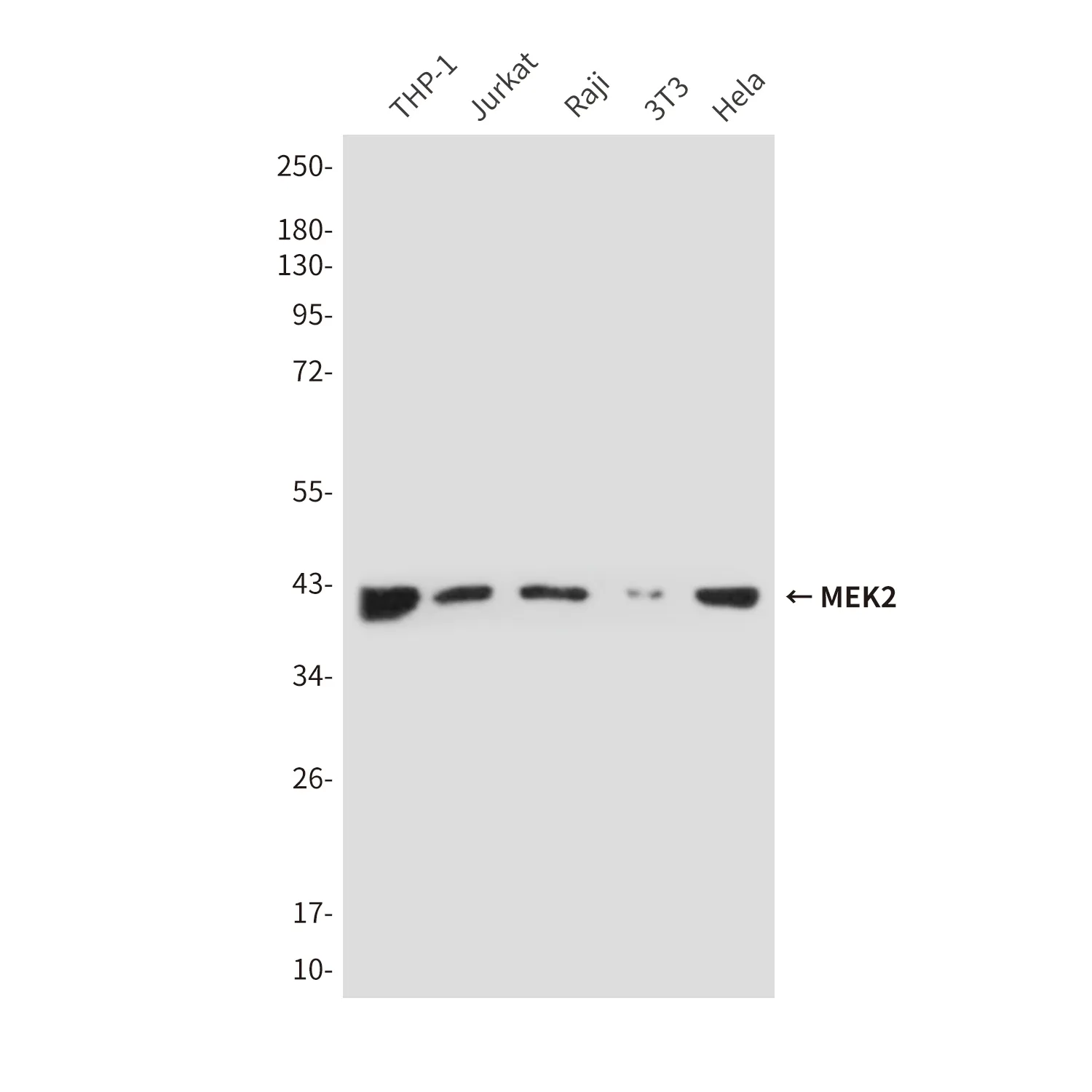
AMPKアルファ1(18A4)ウサギモノクローナル抗体
コンジュゲーション: 非共役
組換えウサギモノクローナル抗体
アプリケーション
反応性
ヒト、マウス、ラット
遺伝子名
PRKAA1
保存
小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。
要約
| 製品名 | AMPKアルファ1(18A4)ウサギモノクローナル抗体 |
| 説明 | 組換えウサギモノクローナル抗体 |
| 宿主 | うさぎ |
| 反応性 | ヒト、マウス、ラット |
| コンジュゲーション | 非共役 |
| 修飾 | 未修正 |
| アイソタイプ | IgG |
| クローン性 | モノクローナル |
| 形態 | 液体 |
| 濃度 | 非共役 |
| 保存 | 小分けして-20℃で保存してください(12ヶ月間有効)。凍結融解サイクルは避けてください。 |
| 配送 | 氷嚢。 |
| バッファー | ウサギIgG(リン酸緩衝生理食塩水、pH 7.4、150mM NaCl、0.02%新型保存料N、50%グリセロール含有)。短期保存は+4℃、長期保存は-20℃で保存してください。凍結融解サイクルは避けてください。 |
| 精製 | アフィニティー精製 |
抗原情報
| 遺伝子名 | PRKAA1 |
| 別名 | AMP-activate kinase alpha 1 subunit; 5'-AMP-activated protein kinase catalytic subunit alpha-1; AAPK1; AMPK; SNF1A; HMGCR kinase; PRKAA1; cb116; |
| 遺伝子ID | 5562 |
| SwissProt ID | Q13131 |
| 免疫原 | ヒトAMPKアルファ1の合成ペプチド |
アプリケーション
| アプリケーション | WB,IHC,ICC/IF,FC,IP |
| 希釈倍率 | WB 1:500-1:2000,IHC 1:100-1:200,ICC/IF 1:100-1:200,FC 1:100-1:200,IP 1:50-1:100 |
| 分子量 | 64kDa |
研究分野
| Regulation of autophagy;mTOR;Insulin_Receptor;Adipocytokine;Hypertrophic cardiomyopathy (HCM) |
背景
| AMPKA1はCAMKLファミリーのタンパク質キナーゼであり、AMP/ATPと細胞内Ca(2+)レベルのバランスに応じて細胞および生物のエネルギーバランスを制御する上で中心的な役割を果たします。AMP活性化タンパク質キナーゼ(AMPK)の触媒サブユニットは、細胞のエネルギー代謝を制御する上で重要な役割を果たすエネルギーセンサータンパク質キナーゼです。細胞内ATPレベルの減少に応答して、AMPKはエネルギー産生経路を活性化し、エネルギー消費プロセスを阻害します。つまり、タンパク質、炭水化物、脂質の生合成、および細胞の成長と増殖を阻害します。AMPKは、代謝酵素の直接リン酸化を介して、および転写制御因子のリン酸化を介してより長期的な効果によって作用します。また、アクチン細胞骨格をリモデリングすることにより、おそらく間接的にミオシンを活性化することにより、細胞極性の調節因子としても機能します。 ACACA、ACACB、GYS1、HMGCR、LIPEなどの脂質代謝酵素をリン酸化・不活性化することで脂質合成を制御します。また、アセチルCoAカルボキシラーゼ(ACACAおよびACACB)とホルモン感受性リパーゼ(LIPE)をそれぞれリン酸化することで脂肪酸とコレステロールの合成を制御します。IRS1、PFKFB2、PFKFB3をリン酸化することでインスリンシグナル伝達と解糖を制御します。AMPKは、グルコーストランスポーターSLC2A4/GLUT4の細胞膜への移行を促進することで筋肉におけるグルコースの取り込みを促進します。これはおそらくTBC1D4/AS160のリン酸化を介していると考えられます。 CRTC2/TORC2、FOXO3、ヒストンH2B、HDAC5、MEF2C、MLXIPL/ChREBP、EP300、HNF4A、p53/TP53、SREBF1、SREBF2、PPARGC1Aといったエネルギー代謝に関与する転写制御因子をリン酸化することにより、転写とクロマチン構造を制御します。肝臓において、CRTC2/TORC2をリン酸化することで、CRTC2/TORC2を細胞質内に隔離し、グルコース恒常性の重要な制御因子として機能します。ストレス応答として、ヒストンH2Bの「Ser-36」(H2BS36ph)をリン酸化することで、転写を促進します。 TSC2、RPTOR、およびATG1/ULK1をリン酸化することで、細胞の成長と増殖の主要な制御因子として機能します。栄養制限に応答して、mTORC1複合体のRPTOR構成要素をリン酸化することにより、およびTSC2をリン酸化して活性化することにより、mTORC1複合体を負に制御します。栄養制限に応答して、ATG1/ULK1をリン酸化して活性化することにより、オートファジーを促進します。その過程で、WDR45も活性化します(PubMed:28561066)。栄養制限に応答して、転写因子FOXO3をリン酸化して、FOXO3のミトコンドリアへの輸入を促進します(類似性による)。AMPKは、CRY1のリン酸化を媒介して不安定化させることで、概日リズムの制御因子としても機能します。CTNNB1をリン酸化してWntシグナル伝達経路を制御し、安定化させる可能性があります。タウタンパク質キナーゼ活性も有する。アミロイドβA4タンパク質(APP)への曝露に反応し、CAMKK2によって活性化され、MAPT/TAUのリン酸化を引き起こす。しかし、このデータの生体内における関連性は未だ不明である。CFTR、EEF2K、KLC1、NOS3、SLC12A1もリン酸化される。 |